
This is a revision in progress of the Nucleolus page



The nucleolus (/njuːˈkliːələs/ or /ˌnjuːkliˈoʊləs/, plural nucleoli /njuːˈkliːəlaɪ/ or /ˌnjuːkliˈoʊlaɪ/ is the largest structure in the nucleus of eukaryotic cells.[1] Nucleoli are made of proteins, DNA and RNA and form around specific chromosomal regions called nucleolar organizing regions. As early as 2010, the authors of a review wrote:
"The nucleolus has been recognized as a dynamic organelle with many more functions than one could imagine."[2]
It is best known as the site of ribosome biogenesis. Nucleoli also participate in the formation of signal recognition particles and play a role in the cell's response to stress.[3] Nucleoli are also involved in cell cycle control,[4] organization of the genome[1] and many other functions. Malfunction of nucleoli can be the cause of several human conditions called "nucleolopathies"[5] and the nucleolus is being investigated as a target for cancer chemotherapy.[6][7]
History edit

tripartite organization
granular component
The nucleolus was identified by simple microscopy in the decade of the 1830s.[8] Until 1964, little was known about the function of the nucleolus. During that year, a study[9] of nucleoli by John Gurdon and Donald Brown in the African clawed frog Xenopus laevis generated increasing interest in the function and detailed structure of the nucleolus. They found that 25% of the frog eggs had no nucleolus and that such eggs were not capable of life. Half of the eggs had one nucleolus and 25% had two. They concluded that the nucleolus had a function necessary for life. In 1966 Max L. Birnstiel and collaborators showed via nucleic acid hybridization experiments that DNA within nucleoli code for ribosomal RNA.[10][11]
Structure edit
Three major components of the nucleolus are recognized: the fibrillar center (FC), the dense fibrillar component (DFC), and the granular component (GC).[1] Transcription of the rDNA occurs in the FC.[12] The DFC contains the protein fibrillarin,[12] which is important in rRNA processing. The GC contains the protein nucleophosmin,[12] (B23 in the external image) which is also involved in ribosome biogenesis. The structure of the nucleous is dependent on ribosomal proteins, most notably RPL5 and RPL11.[13] There is evidence that Alu element RNA and nucleolar proteins form a nucleolar scaffold.[14]
It has been proposed that the tripartite organization is only observed in higher eukaryotes and that it evolved from a bipartite organization with the transition from anamniotes to amniotes. Reflecting the substantial increase in the DNA intergenic region, an original fibrillar component would have separated into the FC and the DFC.[15]
Another structure identified within many nucleoli (particularly in plants) is a clear area in the center of the structure referred to as a nucleolar vacuole.[16] Nucleoli of various plant species have been shown to have very high concentrations of iron[17] in contrast to human and animal cell nucleoli.
The nucleolus ultrastructure can be visualized through an electron microscope, while the organization and dynamics can be studied through fluorescent protein tagging and fluorescent recovery after photobleaching (FRAP). Antibodies against the PAF49 protein can also be used as a marker for the nucleolus in immunofluorescence experiments.[18]
Functions edit
The nucleolus is far more than a factory for ribosome assembly, and doesn't even complete that job, since many steps in ribosome biogenesis occur outside the nucleolus.
Ribosomes and translation edit
In bacteria it is well established that transcription and translation are coupled. That is, translation occurs while the mRNA is being synthesized. It has been shown that ribosomes actually cooperate with RNA polymerase during transcription.[19] The coupling is manifest in the appearance of an "expressome", a complex in which a ribosome and RNA polymerase are in physical contact and together cover approximately 30 bases of the mRNA.[20] This appears to be the case in archaea also.[21]
In spite of reports starting in the 1950s that translation occurred in eukaryotic nuclei,[22][23][24] the reports were not viewed as compelling for various reasons in 2003.[25][26] More recently, using more modern techniques, 3 different laboratories have found evidence for translation in the nucleus.[27] Evidence for mature 80S ribosomes in the nucleolus has been reported[28] and actual translation was observed in the nucleolus.[28][29][30]
quality control/proteasome "Yet, upon genotoxic stress or inhibition of transcription, ATR-mediated PML phosphorylation relocalizes PML to nucleolar caps, where PML was proposed to sequester the p53 ubiquitin ligase MDM2" "The most abundant (but perhaps least studied) isoform, PML-I, harbors an exonuclease-III domain, that targets PML to nucleolar caps in stressed or senescent cells (Condemine et al. 2007)."[31] Promyelocytic leukemia protein#Function
"We show that contrary to prevailing notion, a group of nucleoplasmic proteins, mostly RNA binding proteins, relocalized from the nucleoplasm to a specific nucleolar cap during transcriptional inhibition. For instance, an exclusively nucleoplasmic protein, the splicing factor PSF, localized to nucleolar caps under these conditions. This structure also contained pre-rRNA transcripts, but other caps contained either nucleolar proteins, PML, or Cajal body proteins and in addition nucleolar or Cajal body RNAs. In contrast to the capping of the nucleoplasmic components, nucleolar granular component proteins dispersed into the nucleoplasm, although at least two (p14/ARF and MRP RNA) were retained in the central body. The nucleolar caps are dynamic structures as determined using photobleaching and require energy for their formation. These findings demonstrate that the process of nucleolar segregation and capping involves energy-dependent repositioning of nuclear proteins and RNAs and emphasize the dynamic characteristics of nuclear domain formation in response to cellular stress."[32]
SRP biogenesis edit
are found in the nucleolus
"Subsequently, the link between the nucleolus and signal recognition particle biosynthesis was made in yeast (Ciufo and Brown 2000; Grosshans et al. 2001), and confirmed and extended in mammalian cells (Politz et al. 2000) (Fig. 4) and in Xenopus oocytes (Sommerville et al. 2005). A speculative essay suggested how a common nucleolar assembly site might have co-evolved for the four translational ribonucleoproteins (the two ribosomal subunits, the 5S rRNA–protein complex, and the SRP) (Pederson and Politz 2000). Notwithstanding how it came about in the evolution of eukaryotes, the fact that the nucleolus is a site of more than ribosome synthesis had clearly arrived once its role in the biosynthesis of the SRP had been discovered."[8] The nucleolus and signal recognition particle biosynthesis.[8]
Stress response edit


"Stresses of all kinds affect ribosome activity (e.g. reviewed in (113)), the production of ribosomes, and consequently the nucleolus itself. Responses to stress (e.g. reviewed in (114)) may sometimes involve gross alterations to nucleolar structure (e.g. (115))." This is ref 115[33]
"Forcing nucleolar disruption by anti-upstream binding factor (UBF) microinjection (in the absence of DNA damage) also causes p53 stabilization. We propose that the nucleolus is a stress sensor responsible for maintenance of low levels of p53, which are automatically elevated as soon as nucleolar function is impaired in response to stress. Our model integrates all known p53-inducing agents and also explains cell cycle-related variations in p53 levels which correlate with established phases of nucleolar assembly/disassembly through the cell cycle." from abstract of[33]
"In summary, three novel ribotoxic stress stimuli were revealed with a potential to be used in nucleolus-focused anticancer therapy."[34]
"Beyond its primary role in ribosome biosynthesis, the nucleolus is also involved in the regulation of cell cycle progression and stress signaling [3], [4], [5]."[34] This is 4[35]
Abstract "All organisms sense and respond to conditions that stress their homeostasis by downregulating the synthesis of rRNA and ribosome biogenesis, thus designating the nucleolus as the central hub in coordinating the cellular stress response. One of the most intriguing roles of the nucleolus, long regarded as a mere ribosome-producing factory, is its participation in monitoring cellular stress signals and transmitting them to the RNA polymerase I (Pol I) transcription machinery. As rRNA synthesis is a most energy-consuming process, switching off transcription of rRNA genes is an effective way of saving the energy required to maintain cellular homeostasis during acute stress. The Pol I transcription machinery is the key convergence point that collects and integrates a vast array of information from cellular signaling cascades to regulate ribosome production which, in turn, guides cell growth and proliferation. This review focuses on the mechanisms that link cell physiology to rDNA silencing, a prerequisite for nucleolar integrity and cell survival."[35]
"The nucleolus controls ribosome biogenesis and its perturbation induces nucleolar stress that inhibits cell cycle progression and activates checkpoint responses. Here, we investigate the roles of ribosomal RNA processing protein, RRP15, in nucleolar formation, ribosome biogenesis, cell cycle progression and checkpoint control in human cells. RRP15 is localized in the nucleolus and required for nucleolar formation. Perturbation of RRP15 induces nucleolar stress that activates RPL5/RPL11/5S rRNA (RP)-Mdm2-p53 axis checkpoint response and arrests cells at G1-G1/S in p53-proficient non-transformed RPE1 cells"[36]
Abstract "The seminal identification of ribosomal protein binding to MDM2,[a] the negative regulator of p53, has evolved into a paradigm for ribosomal protein–MDM2–p53 signaling that extends into processes as diverse as energy metabolism to proliferation. The central core of signaling occurs when perturbations to rRNA synthesis, processing, and assembly modulate the rate of ribosome biogenesis, signaling a nucleolar stress response to p53. This has led to identification of a number of disease pathologies related to ribosomal protein dysfunction that are manifested as developmental disorders or cancer." [37]
from abstract "Furthermore, both phosphorylation and degradation of PICT-1 released RPL11 from the nucleolus to the nucleoplasm, increased binding of RPL11 to MDM2, and promoted p53 accumulation and apoptosis in an ATM-dependent manner after DNA damage. These data indicate that PICT-1 is a major nucleolar sensor of the DNA damage repair response and an important upstream regulator of p53 via the RPL11-MDM2-p53 pathway."[38]
from abstract "In this review, we focus on how two nuclear organelles, the nucleolus and the Cajal body, respond to stress. The nucleolus senses stress and is a central hub for coordinating the stress response. We review nucleolar function in the stress-induced regulation of p53 and the specific changes in nucleolar morphology and composition that occur upon stress. Crosstalk between nucleoli and CBs is also discussed in the context of stress responses."[39]
"In addition, different types of stress that affect CB integrity, such as the inhibition of transcription and/or snRNA processing, induce the redistribution of coilin to the nucleolus (Figure 1C and Table 1)."[39]
from abstract"The nucleoplasmic translocation of nucleolar protein nucleophosmin (NPM1) is a hallmark of nucleolar stress; however, the causes of this translocation and its connection to p53 activation are unclear. Using single live-cell imaging and the redox biosensors, we demonstrate that nucleolar oxidation is a general response to various cellular stresses. During nucleolar oxidation, NPM1 undergoes S-glutathionylation on cysteine 275, which triggers the dissociation of NPM1 from nucleolar nucleic acids. The C275S mutant NPM1, unable to be glutathionylated, remains in the nucleolus under nucleolar stress. Compared with wild-type NPM1 that can disrupt the p53–HDM2 interaction, the C275S mutant greatly compromises the activation of p53, highlighting that nucleoplasmic translocation of NPM1 is a prerequisite for stress-induced activation of p53."[40]
from abstract "Forcing nucleolar disruption by anti-upstream binding factor (UBF) microinjection (in the absence of DNA damage) also causes p53 stabilization. We propose that the nucleolus is a stress sensor responsible for maintenance of low levels of p53, which are automatically elevated as soon as nucleolar function is impaired in response to stress."[41]
Cell cycle control edit
The nucleolus functions in cell cycle control, reviewed in.[42]
"Several proteins are known to localize to the nucleolus in a cell cycle-specific manner, including several that are associated with human disease (26)."[1]
"nucleolar structure changes in response to both environmental conditions and the cell cycle (18, 26)"[1]
"Such structural alterations, as well as alterations in the numbers of rDNA repeats, would relieve or exacerbate the retention of proteins sequestered in the nucleolus as a result of changes in the spatial clustering of binding sites."[1]
organizer of genome structure edit
"There is growing evidence that the genome takes on a specific structural arrangement within the nucleus. In human cells, different chromosomes are found to occupy chromosome “territories”, which have different positions in different cell types (28, 29)."[1]
"Computational-based approaches, utilising proximity-ligation data and biophysical characteristics, have been taken to model global genome structure (e.g. (38, 40, 43, 44)). Interestingly, few restraints are required to impart a crude order on in silico polymer-based reconstructions of the budding yeast nucleus (43, 44). However, one restraint that is required is the positioning of the nucleolus opposite to the spindle pole body (43), suggesting the nucleolus is a significant landmark for spatial organization of the genome."[1]
"However, these differences in the linear organization of the 5S genes between species belie commonalities in their spatial localization. For example, in mice ectopic 5S rDNA gene sequences have been shown to promote nucleolar localisation (71). Similarly, in humans, one of the transcribed, linear clusters of 5S genes on chromosome I was shown to localize to a perinucleolar compartment (72). Moreover, the linearly dispersed 5S genes in many other eukaryotes have been shown to co-localize with nucleoli in three dimensions (73), suggesting that there are benefits to co-localizing the 5S genes with the other ribosomal genes. This is strong evidence for the nucleolus playing a direct role in the spatial organisation of the nucleus."[1]
"In S. cerevisiae both microscopy and crosslinking proximity analysis show that tRNA genes cluster together and co-localize with the nucleolus (37, 42, 79–81)."[1]
"Little is known about the three-dimensional organization of tRNA genes in most eukaryotes, however, and whether they co-localize with nucleoli."[1]
"The nucleolus appears to influence the chromatin structure of the DNA that surrounds it. In metazoans the nucleolus is commonly observed to be surrounded by shell of late-replicating heterochromatin."[1]
"Furthermore, nucleolar association seems to be an important factor to maintain the heterochromatic state of the inactive human X chromosome (95), with the Barr body originally being known as the “nucleolar satellite” (96)."[1] This is ref 96[43]
Virus replication edit
"nucleolar localization of viral proteins involved in viral replication, including HIV, appears to be necessary for replication (19)."[1]
"From the integral nature of the nucleolus to many RNA processing and RNP assembly pathways, it is not surprising that many animal and plant viruses exploit the nucleolus in production and transport of viral RNPs. The involvement of the nucleolus in infection cycles of animal and human viruses is well established (Hiscox, 2007; Hiscox et al., 2010; Taliansky et al., 2011). In plants, a growing number of viruses show some interaction with the nucleolus (and other nuclear bodies such as CBs) and roles for the nucleolus and nucleolar proteins are now emerging (Taliansky et al., 2011). For example, plant viruses can recruit nucleolar proteins for assembly of viral RNP particles, virus replication and movement, and to counteract host-viral defense systems. In the best-studied system to date, the ORF3 long-distance movement protein of Groundnut rosette virus trafficks to the nucleolus via CBs, causing reorganization of CBs into multiple CB-like structures that fuse with the nucleolus. ORF3 then recruits fibrillarin (an abundant nucleolar RNA binding protein, known to be an RNA methylase) for assembly of cytoplasmic infectious viral particles (Kim et al., 2007a, 2007b; Canetta et al., 2008). The nucleolar localization of ORF3 is essential for systemic infection. Viral proteins from other plant viruses also target the nucleolus and the function of this localization and their interactions with nucleolar proteins are being established. For example, the coat protein and coat protein read-through proteins of Potato leaf roll virus (PLRV) are targeted to the nucleolus (Haupt et al., 2005) and systemic infection of PLRV is inhibited in fibrillarin-silenced plants, suggesting that fibrillarin is also involved in long-distance movement of PLRV."[42]
protein sequestration edit
"Strikingly, several noncoding RNA transcripts from the rDNA IGS appear to bind and sequester proteins in the nucleolus, and are regulated by stress (27)."[1] This is 27[44]
" Recent study of the IGS has contradicted the dogma that these spacers are merely inert regions of the genome, instead suggesting they are biologically significant, complex and plurifunctional transcriptional units that appear central to proper cellular functioning. Through the timely induction of various ribosomal IGS noncoding RNA (IGS RNA) transcripts, the cell is capable of both regulating rRNA synthesis and sequestering large numbers of proteins, thereby modulating essential molecular networks." from abstract of [44]
" we performed a genome-wide microscopy-based screening for proteins that form nuclear foci and characterized their localizations using markers of known nuclear bodies. In total, we identified 325 proteins localized to distinct nuclear bodies, including nucleoli (148), promyelocytic leukemia nuclear bodies (38), nuclear speckles (27), paraspeckles (24), Cajal bodies (17), Sam68 nuclear bodies (5), Polycomb bodies (2), and uncharacterized nuclear bodies (64). Functional validation revealed several proteins potentially involved in the assembly of Cajal bodies and paraspeckles. Together, these data establish the first atlas of human proteins in different nuclear bodies and provide key information for research on nuclear bodies."[45] Comparison of our study with datasets on nuclear subcompartments[45]
"From the initial ∼700 human proteins identified in the previous iteration of the NOPdb, we have now identified over 50 000 peptides contained in over 4500 human proteins from purified nucleoli, providing enhanced coverage of the nucleolar proteome."[46]
"Protein Mobility For many years cell biologists visualized fixed cells and thus had a tendency to regard the structures seen as stationary and long lived. Live-cell imaging studies, however, have revealed a much more dynamic picture. Studies using fluorescence recovery after photobleaching of nucleolar and nuclear proteins fused to GFP have shown that virtually all nucleolar and nuclear proteins are in constant flux, exchanging between the nucleolus and cytoplasm. The mean nucleolar residence time of even well-characterized nucleolar proteins is only a few tens of seconds (Phair and Misteli, 2000; Chen and Huang, 2001; Olson and Dundr, 2005). The distinction between nuclear and nucleolar proteins is that nucleolar proteins spend a greater proportion of their time in the nucleolus."[42]
Telomeres and heterochromatin edit

"we show that TRF2 normally associates with the nucleolus and rDNA. However, when telomeres are crippled by dnTRF2 or RNAi knockdown of TRF2, gross nucleolar and chromosomal changes occur. We used the controllable dnTRF2 system to precisely dissect the timing and progression of nucleolar and chromosomal instability induced by telomere dysfunction, demonstrating that nucleolar changes precede the DNA damage and morphological changes that occur at acrocentric short arms. The rDNA repeat arrays on the short arms decondense, and are coated by RNA polymerase I transcription binding factor UBF, physically linking acrocentrics to one another as they become fusogenic. These results highlight the importance of telomere function in nucleolar stability and structural integrity of acrocentric chromosomes, particularly the rDNA arrays. Telomeric stress is widely accepted to cause DNA damage at chromosome ends, but our findings suggest that it also disrupts chromosome structure beyond the telomere region, specifically within the rDNA arrays located on acrocentric chromosomes. These results have relevance for Robertsonian translocation formation in humans and mechanisms by which acrocentric-acrocentric fusions are promoted by DNA damage and repair."[47]
"A shell of pericentric heterochromatin typically surrounds nucleoli. Using deep sequencing two independent studies recently provided a detailed analysis of DNA attached to isolated nucleoli, which were termed nucleolus-associated chromatin domains (NADs) [15 ,16]."[48]
from 16 abstract "The data show that specific sequences from most human chromosomes, in addition to the rDNA repeat units, associate with nucleoli in a reproducible and heritable manner. NADs have in common a high density of AT-rich sequence elements, low gene density and a statistically significant enrichment in transcriptionally repressed genes."[49]
from 15 abstract "revealed the pattern of nucleolus-associated chromatin domains (NADs) in the linear human genome and identified different gene families and certain satellite repeats as the major building blocks of NADs, which constitute about 4% of the genome. Bioinformatic evaluation showed that NAD–localized genes take part in specific biological processes, like the response to other organisms, odor perception, and tissue development. 3D FISH and immunofluorescence experiments illustrated the spatial distribution of NAD–specific chromatin within interphase nuclei and its alteration upon transcriptional changes"[50]
In 2 casees, the Barr body and KCNQ1 imprinted gene locus, the inteactions of heterochromatin with the nucleolar periphery is dependent upon RNA (xist and KCNQ1OT1)[48] In other cases, various proteins have been implicated.[48]
Disease associations edit
Sequestration of proteins edit
In addition to its role in ribosomal biogenesis, the nucleolus is known to capture and immobilize proteins, a process known as nucleolar detention. Proteins that are detained in the nucleolus are unable to diffuse and to interact with their binding partners. Targets of this post-translational regulatory mechanism include VHL, PML, MDM2, POLD1, RelA, HAND1 and hTERT, among many others. It is now known that long noncoding RNAs originating from intergenic regions of the nucleolus are responsible for this phenomenon.[51]
| Cell biology | |
|---|---|
| Animal cell diagram | |
 Components of a typical animal cell:
|
Intanucleolar bodies edit
"Using high resolution immunoelectron microscopy (I-EM), we provide evidence that CNoBs (CRM1-Nucleolar bodies) and INBs (Intranucleolar bodies) are distinct genuine nucleolar structures in untreated HeLa cells."[52]
"Comparing CNoBs, INBs and CBs Previous IF studies have reported the presence of CRM1 in CBs.30,31 However, none of the vitreous CRM1-positive bodies that we identified by I-EM resembled CBs. In addition, visual inspection of the CBs, ultrastructurally characterized by 40–60 nm thick coiled threads on the same thin-sections, revealed that they were not labeled (Fig. S2). Since this discrepancy could result from the different antibodies used in these studies, we labeled thin-sections with a distinct anti-CRM1 antibody that again decorated the same vitreous bodies, but not the CBs (Fig. S2B). On the other hand, the CB marker coilin,32 was detected in INBs,16 and in intranucleolar CBs in breast cancer cell lines,33,34 or upon okadaic acid treatment.35 Using an anti-coilin antibody on HeLa cell thin-sections, heavily labeled CBs (297 +/− 87 gold particles/µm2, n = 16) were found within the nucleoplasm whereas the nucleolar-associated and nucleoplasmic vitreous CNoBs remained unlabelled (Fig. S3A and A′), in agreement with previous IF observations.15 In the same cell sample, the INBs recognized by their ultrastructural features (n = 5) were also unlabeled (Fig. S3B). Overall, as illustrated in Figure 3 and Figures S2 and S3, CBs, INBs and CNoBs are so dissimilar in size and so distinct ultrastructurally in HeLa cells that they cannot be confounded; confirming that the CRM1-positive nucleoplasmic bodies seen under the EM are CNoBs and the SUMO-labeled intranucleolar bodies are INBs, with no contribution of the CBs."[52] Characterization of INBs by I-EM localization of SUMO-2/3 and SUMO1>[52]
"the de novo formation of a large subnucleolar structure, termed the detention center (DC). The DC is a spatially and dynamically distinct region, characterized by an 8-anilino-1-naphthalenesulfonate–positive hydrophobic signature. Its formation is accompanied by redistribution of nucleolar factors and arrest in ribosomal biogenesis. Silencing of regulatory IGS lncRNA prevents the creation of this structure and allows the nucleolus to retain its tripartite organization and transcriptional activity."[53] The nucleolus responds structurally to environmental stimuli.[53]
The nucleolus is reorganized around the detention center.[53] Model: functional remodeling of the nucleolus.[53]
"Here, we describe the localisation of 21 protein factors to an intranucleolar region different to these main subcompartments, called the intranucleolar body (INB)." " Increase in the size and number of INBs is promoted by specific types of DNA damage and depends on the functional integrity of the nucleolus. INBs are abundant in nucleoli of unstressed cells during S phase and localise in close proximity to rDNA with heterochromatic features. The data suggest the INB is linked with regulation of rDNA transcription and/or maintenance of rDNA."[54]
"The observation that the INB is mostly abundant in cells during S phase, localises in close proximity to rDNA and is promoted by specific DNA damaging agents might indicate that, at least in part, its components are linked to the maintenance of rDNA repeats."[54]
"A new intranucleolar body (INB) has recently been described, based on colocalization of about 20 well-characterized components (Hutten et al., 2011). The composition of INBs seems distinctly different from CBs; in particular they contain small ubiquitin-like modifier, a peptide modifier, probably conjugated to other substrate proteins. The INBs seem to have an involvement in DNA damage response, since treatments that caused DNA damage induced INBs. The INBs are completely enveloped in the nucleolus, and may be contained in or overlap with a central region of the nucleolus often called the nucleolar cavity, a structure that is particularly prominent in many plant cells and that has been shown to swell and contract dynamically."[42]
"CBs are intimately linked with the nucleolus, both on physical and functional levels. CBs were originally called “nucleolar accessory bodies” because of their close association with nucleoli in neurons (Gall, 2000; Lafarga et al., 2009). CBs are even sometimes found within the nucleolus and a budding yeast counterpart of the CB, termed the “nucleolar body,” is part of the nucleolus (Cioce and Lamond, 2005; Verheggen et al., 2002). In addition, CBs are involved in the maturation of snoRNPs, which are subsequently transported to the nucleoli, where they participate in rRNA processing. In general, both nucleoli and CBs are involved in the production of non-poly(A)-tailed RNAs that are tightly connected to cell growth, including histone mRNAs, snRNAs, and snoRNAs in CBs, and rRNAs in nucleoli. Consistent with this, both structures are prominent in cells that are transcriptionally and metabolically active, such as neuronal and cancer cells (Berciano et al., 2007)."[39]
Assembly and disassembly (and maybe remodeling, including detention center) edit
"The assembly of the nucleoli during telophase/early G1 depends on pre-existing machineries inactivated during prophase (the transcription machinery and RNP processing complexes) and on partially processed 45S rRNAs inherited throughout mitosis. In telophase, the 45S rRNAs nucleate the prenucleolar bodies and order the dynamics of nucleolar assembly. The assembly/disassembly processes of the nucleolus depend on the equilibrium between phosphorylation/dephosphorylation of the transcription machinery and on the RNP processing complexes under the control of the CDK1-cyclin B kinase and PP1 phosphatases. The dynamics of assembly/disassembly of the nucleolus is time and space regulated."[55]
Nucleolinus edit
"The nucleolinus is a little-known cellular structure, discovered over 150 years ago" "Analysis of nucleolinar RNA and protein revealed a set of molecules that overlaps with but is nevertheless distinct from the nucleolus. The proteins identified were primarily ones involved in nucleic acid metabolism and cell cycle regulation. Monoclonal antibodies generated against isolated nucleolini revealed centrosomal forerunners in the oocyte cytoplasm. Finally, induction of damage to the nucleolinus by laser microsurgery altered the trafficking of α- and γ-tubulin after fertilization. These observations strongly support a role for the nucleolinus in cell division and represent our first clues regarding mechanism."[56] Isolation of nucleolini.[56]
"we isolated nucleolini, extracted their RNA, and prepared an in situ riboprobe (NLi-1), which is associated specifically with the nucleolinus, confirming its unique composition. Other in situ observations revealed a NLi-1 and nucleolinar association with the developing spindle and centrosomes."[57] DIC image of the nucleolinus within an unactivated Spisula oocyte[57] Spatial relationship of the nucleolinus to the developing spindle and pole in a living oocyte and movie link.[57] By 12 min postactivation (C), the entire cell division apparatus is associated with an NLi-1 “matrix.”[57]
" This brief article relates most of what we know about the nucleolinus and where to find more information."[58]
"The nucleolinus is a nuclear subcompartment long ago posited to play a role in cell division. In a recent study using surf clam oocytes, cytoplasmic foci containing a nucleolinar protein were shown to later recruit γ-tubulin, identifying them as centrosomal precursors.1 We now demonstrate the presence of structural RNAs from the nucleolinus in these procentrosomes. They include the well-known but poorly understood rRNA-transcribed spacer regions."[59]
See also edit
Gallery edit

fBALM DNA super-resolution microscopy of HeLa cell nucleus 
A new stain shows both nuclei and nucleoli in melanoma cells. 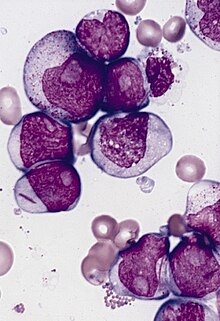
Cells from a patient with acute myeloblastic leukemia with maturation showing several blasts with prominent nucleoli.



- image with links to movies
Notes edit
- ^ MDM2 and hDM2 refer to the same protein. Think of M for Mouse and h for human.
References edit
- ^ a b c d e f g h i j k l m n o p O'Sullivan JM, Pai DA, Cridge AG, Engelke DR, Ganley AR (2013). "The nucleolus: a raft adrift in the nuclear sea or the keystone in nuclear structure?". Biomolecular Concepts. 4 (3): 277–86. doi:10.1515/bmc-2012-0043. PMC 5100006. PMID 25436580.
- ^ Cisterna B, Biggiogera M (2010). "Ribosome biogenesis: from structure to dynamics". International Review of Cell and Molecular Biology. 284: 67–111. doi:10.1016/S1937-6448(10)84002-X. PMID 20875629.
- ^ Olsen, Mark OJ (December 2010). "Nucleolus: Structure and Function". ELS. doi:10.1002/9780470015902.a0005975.pub2. ISBN 0470016175. Retrieved October 17, 2014.
- ^ Tsai RY, Pederson T (2014). "Connecting the nucleolus to the cell cycle and human disease". FASEB Journal : Official Publication of the Federation of American Societies for Experimental Biology. 28 (8): 3290–6. doi:10.1096/fj.14-254680. PMID 24790035.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Hetman M (2014). "Role of the nucleolus in human diseases. Preface". Biochimica Et Biophysica Acta. 1842 (6): 757. doi:10.1016/j.bbadis.2014.03.004. PMID 24631655.
- ^ Quin JE, Devlin JR, Cameron D, Hannan KM, Pearson RB, Hannan RD (2014). "Targeting the nucleolus for cancer intervention". Biochimica Et Biophysica Acta. 1842 (6): 802–16. doi:10.1016/j.bbadis.2013.12.009. PMID 24389329.
- ^ Woods SJ, Hannan KM, Pearson RB, Hannan RD (2015). "The nucleolus as a fundamental regulator of the p53 response and a new target for cancer therapy". Biochimica Et Biophysica Acta. 1849 (7): 821–9. doi:10.1016/j.bbagrm.2014.10.007. PMID 25464032.
- ^ a b c d Pederson T (2011). "The nucleolus". Cold Spring Harbor Perspectives in Biology. 3 (3). doi:10.1101/cshperspect.a000638. PMC 3039934. PMID 21106648.
- ^ BROWN DD, GURDON JB (1964). "ABSENCE OF RIBOSOMAL RNA SYNTHESIS IN THE ANUCLEOLATE MUTANT OF XENOPUS LAEVIS" (PDF). Proceedings of the National Academy of Sciences of the United States of America. 51: 139–46. PMC 300879. PMID 14106673.
- ^ Birnstiel ML, Wallace H, Sirlin JL, Fischberg M (1966). "Localization of the ribosomal DNA complements in the nucleolar organizer region of Xenopus laevis". National Cancer Institute Monograph. 23: 431–47. PMID 5963987.
- ^ Wallace H, Birnstiel ML (1966). "Ribosomal cistrons and the nucleolar organizer". Biochimica Et Biophysica Acta. 114 (2): 296–310. PMID 5943882.
- ^ a b c d Sirri V, Urcuqui-Inchima S, Roussel P, Hernandez-Verdun D (2008). "Nucleolus: the fascinating nuclear body". Histochemistry and Cell Biology. 129 (1): 13–31. doi:10.1007/s00418-007-0359-6. PMC 2137947. PMID 18046571.
- ^ Nicolas E, Parisot P, Pinto-Monteiro C, de Walque R, De Vleeschouwer C, Lafontaine DL (2016). "Involvement of human ribosomal proteins in nucleolar structure and p53-dependent nucleolar stress". Nature Communications. 7: 11390. doi:10.1038/ncomms11390. PMC 4897761. PMID 27265389.
- ^ Carmo-Fonseca M (2015). "Assembly of the nucleolus: in need of revision". The EMBO Journal. 34 (22): 2731–2. doi:10.15252/embj.201593185. PMC 4682648. PMID 26471727.
- ^ Thiry M, Lafontaine DL (2005). "Birth of a nucleolus: the evolution of nucleolar compartments". Trends Cell Biol. 15 (4): 194–9. doi:10.1016/j.tcb.2005.02.007. PMID 15817375. as PDF
- ^ Beven AF, Lee R, Razaz M, Leader DJ, Brown JW, Shaw PJ (1 June 1996). "The organization of ribosomal RNA processing correlates with the distribution of nucleolar snRNAs". J. Cell. Sci. 109 (6): 1241–51. PMID 8799814.
- ^ Roschzttardtz H, Grillet L, Isaure MP, et al. (2011). "Plant cell nucleolus as a hot spot for iron". J. Biol. Chem. 286 (32): 27863–6. doi:10.1074/jbc.C111.269720. PMC 3151030. PMID 21719700.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ PAF49 antibody | GeneTex Inc. Genetex.com. Retrieved on 2013-03-03.
- ^ Proshkin, S.; Rahmouni, A.; Mironov, A.; Nudler, E. (2010). "Cooperation Between Translating Ribosomes and RNA Polymerase in Transcription Elongation". Science. 328 (5977): 504–508. doi:10.1126/science.1184939. ISSN 0036-8075.
- ^ Kohler R, Mooney RA, Mills DJ, Landick R, Cramer P (2017). "Architecture of a transcribing-translating expressome". Science (New York, N.Y.). 356 (6334): 194–197. doi:10.1126/science.aal3059. PMID 28408604.
- ^ French SL, Santangelo TJ, Beyer AL, Reeve JN (2007). "Transcription and translation are coupled in Archaea". Molecular Biology and Evolution. 24 (4): 893–5. doi:10.1093/molbev/msm007. PMID 17237472.
- ^ ALLFREY VG, MIRSKY AE (1955). "Protein synthesis in isolated cell nuclei". Nature. 176 (4492): 1042–9. PMID 13272745.
- ^ Allfrey VG (1954). "AMINO ACID INCORPORATION BY ISOLATED THYMUS NUCLEI. I. THE ROLE OF DESOXYRIBONUCLEIC ACID IN PROTEIN SYNTHESIS". Proceedings of the National Academy of Sciences of the United States of America. 40 (10): 881–5. PMC 534190. PMID 16589585.
- ^ Iborra FJ, Jackson DA, Cook PR (2001). "Coupled transcription and translation within nuclei of mammalian cells". Science (New York, N.Y.). 293 (5532): 1139–42. doi:10.1126/science.1061216. PMID 11423616.
- ^ Dahlberg JE, Lund E, Goodwin EB (2003). "Nuclear translation: what is the evidence?". RNA (New York, N.Y.). 9 (1): 1–8. PMC 1370363. PMID 12554869.
- ^ Nathanson L, Xia T, Deutscher MP (2003). "Nuclear protein synthesis: a re-evaluation". RNA (New York, N.Y.). 9 (1): 9–13. PMC 1370364. PMID 12554870.
- ^ Apcher S, Millot G, Daskalogianni C, Scherl A, Manoury B, Fåhraeus R (2013). "Translation of pre-spliced RNAs in the nuclear compartment generates peptides for the MHC class I pathway". Proceedings of the National Academy of Sciences of the United States of America. 110 (44): 17951–6. doi:10.1073/pnas.1309956110. PMC 3816435. PMID 24082107.
- ^ a b Al-Jubran K, Wen J, Abdullahi A, Roy Chaudhury S, Li M, Ramanathan P, Matina A, De S, Piechocki K, Rugjee KN, Brogna S (2013). "Visualization of the joining of ribosomal subunits reveals the presence of 80S ribosomes in the nucleus". RNA (New York, N.Y.). 19 (12): 1669–83. doi:10.1261/rna.038356.113. PMC 3884666. PMID 24129492.
- ^ David A, Dolan BP, Hickman HD, Knowlton JJ, Clavarino G, Pierre P, Bennink JR, Yewdell JW (2012). "Nuclear translation visualized by ribosome-bound nascent chain puromycylation". The Journal of Cell Biology. 197 (1): 45–57. doi:10.1083/jcb.201112145. PMC 3317795. PMID 22472439.
- ^ David A, Yewdell JW (2015). "Applying the ribopuromycylation method to detect nuclear translation". Methods in Molecular Biology (Clifton, N.J.). 1228: 133–42. doi:10.1007/978-1-4939-1680-1_11. PMID 25311127.
- ^ Lallemand-Breitenbach V, de Thé H (2010). "PML nuclear bodies". Cold Spring Harbor Perspectives in Biology. 2 (5): a000661. doi:10.1101/cshperspect.a000661. PMC 2857171. PMID 20452955.
- ^ Shav-Tal, Y. (2005). "Dynamic Sorting of Nuclear Components into Distinct Nucleolar Caps during Transcriptional Inhibition". Molecular Biology of the Cell. 16 (5): 2395–2413. doi:10.1091/mbc.E04-11-0992. ISSN 1059-1524.
- ^ a b Rubbi CP, Milner J (2003). "Disruption of the nucleolus mediates stabilization of p53 in response to DNA damage and other stresses". The EMBO Journal. 22 (22): 6068–77. doi:10.1093/emboj/cdg579. PMC 275437. PMID 14609953.
- ^ a b Lewinska A, Bednarz D, Adamczyk-Grochala J, Wnuk M (2017). "Phytochemical-induced nucleolar stress results in the inhibition of breast cancer cell proliferation". Redox Biology. 12: 469–482. doi:10.1016/j.redox.2017.03.014. PMC 5362140. PMID 28334682.
- ^ a b Grummt I (2013). "The nucleolus—guardian of cellular homeostasis and genome integrity". Chromosoma. 122 (6): 487–97. doi:10.1007/s00412-013-0430-0. PMID 24022641.
- ^ Dong Z, Zhu C, Zhan Q, Jiang W (2017). "The roles of RRP15 in nucleolar formation, ribosome biogenesis and checkpoint control in human cells". Oncotarget. 8 (8): 13240–13252. doi:10.18632/oncotarget.14658. PMC 5355092. PMID 28099941.
- ^ Deisenroth, Chad; Franklin, Derek A.; Zhang, Yanping (2016). "The Evolution of the Ribosomal Protein–MDM2–p53 Pathway". Cold Spring Harbor Perspectives in Medicine. 6 (12): a026138. doi:10.1101/cshperspect.a026138. ISSN 2157-1422.
- ^ Chen H, Han L, Tsai H, Wang Z, Wu Y, Duo Y, Cao W, Chen L, Tan Z, Xu N, Huang X, Zhuang J, Huang L (2016). "PICT-1 is a key nucleolar sensor in DNA damage response signaling that regulates apoptosis through the RPL11-MDM2-p53 pathway". Oncotarget. 7 (50): 83241–83257. doi:10.18632/oncotarget.13082. PMC 5347766. PMID 27829214.
- ^ a b c Boulon S, Westman BJ, Hutten S, Boisvert FM, Lamond AI (2010). "The nucleolus under stress". Molecular Cell. 40 (2): 216–27. doi:10.1016/j.molcel.2010.09.024. PMC 2987465. PMID 20965417.
- ^ Yang K, Wang M, Zhao Y, Sun X, Yang Y, Li X, Zhou A, Chu H, Zhou H, Xu J, Wu M, Yang J, Yi J (2016). "A redox mechanism underlying nucleolar stress sensing by nucleophosmin". Nature Communications. 7: 13599. doi:10.1038/ncomms13599. PMC 5133708. PMID 27886181.
- ^ Rubbi CP, Milner J (2003). "Disruption of the nucleolus mediates stabilization of p53 in response to DNA damage and other stresses". The EMBO Journal. 22 (22): 6068–77. doi:10.1093/emboj/cdg579. PMC 275437. PMID 14609953.
- ^ a b c d Shaw P, Brown J (2012). "Nucleoli: composition, function, and dynamics". Plant Physiology. 158 (1): 44–51. doi:10.1104/pp.111.188052. PMC 3252080. PMID 22082506.
{{cite journal}}: Check|url=value (help) - ^ BARR ML, BERTRAM EG (1949). "A morphological distinction between neurones of the male and female, and the behaviour of the nucleolar satellite during accelerated nucleoprotein synthesis". Nature. 163 (4148): 676. PMID 18120749.
- ^ a b Jacob MD, Audas TE, Mullineux ST, Lee S (2012). "Where no RNA polymerase has gone before: novel functional transcripts derived from the ribosomal intergenic spacer". Nucleus (Austin, Tex.). 3 (4): 315–9. doi:10.4161/nucl.20585. PMID 22688644.
- ^ a b Fong KW, Li Y, Wang W, Ma W, Li K, Qi RZ, Liu D, Songyang Z, Chen J (2013). "Whole-genome screening identifies proteins localized to distinct nuclear bodies". The Journal of Cell Biology. 203 (1): 149–64. doi:10.1083/jcb.201303145. PMC 3798253. PMID 24127217.
- ^ Ahmad Y, Boisvert FM, Gregor P, Cobley A, Lamond AI (2009). "NOPdb: Nucleolar Proteome Database--2008 update". Nucleic Acids Research. 37 (Database issue): D181–4. doi:10.1093/nar/gkn804. PMC 2686505. PMID 18984612.
- ^ Stimpson KM, Sullivan LL, Kuo ME, Sullivan BA (2014). "Nucleolar organization, ribosomal DNA array stability, and acrocentric chromosome integrity are linked to telomere function". Plos One. 9 (3): e92432. doi:10.1371/journal.pone.0092432. PMC 3963894. PMID 24662969.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ a b c Padeken J, Heun P (2014). "Nucleolus and nuclear periphery: velcro for heterochromatin". Current Opinion in Cell Biology. 28: 54–60. doi:10.1016/j.ceb.2014.03.001. PMID 24690547.
- ^ van Koningsbruggen S, Gierlinski M, Schofield P, Martin D, Barton GJ, Ariyurek Y, den Dunnen JT, Lamond AI (2010). "High-resolution whole-genome sequencing reveals that specific chromatin domains from most human chromosomes associate with nucleoli". Molecular Biology of the Cell. 21 (21): 3735–48. doi:10.1091/mbc.E10-06-0508. PMC 2965689. PMID 20826608.
- ^ Németh A, Conesa A, Santoyo-Lopez J, Medina I, Montaner D, Péterfia B, Solovei I, Cremer T, Dopazo J, Längst G (2010). "Initial genomics of the human nucleolus". PLoS Genetics. 6 (3): e1000889. doi:10.1371/journal.pgen.1000889. PMC 2845662. PMID 20361057.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Audas TE, Jacob MD, Lee S (2012). "Immobilization of proteins in the nucleolus by ribosomal intergenic spacer noncoding RNA". Mol Cell. 45 (2): 147–57. doi:10.1016/j.molcel.2011.12.012. PMID 22284675.
- ^ a b c Souquere S, Weil D, Pierron G (2015). "Comparative ultrastructure of CRM1-Nucleolar bodies (CNoBs), Intranucleolar bodies (INBs) and hybrid PML/p62 bodies uncovers new facets of nuclear body dynamic and diversity". Nucleus (Austin, Tex.). 6 (4): 326–38. doi:10.1080/19491034.2015.1082695. PMC 4615761. PMID 26275159.
- ^ a b c d Jacob MD, Audas TE, Uniacke J, Trinkle-Mulcahy L, Lee S (2013). "Environmental cues induce a long noncoding RNA-dependent remodeling of the nucleolus". Molecular Biology of the Cell. 24 (18): 2943–53. doi:10.1091/mbc.E13-04-0223. PMC 3771955. PMID 23904269.
- ^ a b Hutten S, Prescott A, James J, Riesenberg S, Boulon S, Lam YW, Lamond AI (2011). "An intranucleolar body associated with rDNA". Chromosoma. 120 (5): 481–99. doi:10.1007/s00412-011-0327-8. PMC 3232531. PMID 21698343.
- ^ Hernandez-Verdun D (2011). "Assembly and disassembly of the nucleolus during the cell cycle". Nucleus (Austin, Tex.). 2 (3): 189–94. doi:10.4161/nucl.2.3.16246. PMC 3149879. PMID 21818412.
- ^ a b Alliegro MC, Hartson S, Alliegro MA (2012). "Composition and dynamics of the nucleolinus, a link between the nucleolus and cell division apparatus in surf clam (Spisula) oocytes". The Journal of Biological Chemistry. 287 (9): 6702–13. doi:10.1074/jbc.M111.288506. PMC 3307295. PMID 22219192.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ a b c d Alliegro MA, Henry JJ, Alliegro MC (2010). "Rediscovery of the nucleolinus, a dynamic RNA-rich organelle associated with the nucleolus, spindle, and centrosomes". Proceedings of the National Academy of Sciences of the United States of America. 107 (31): 13718–23. doi:10.1073/pnas.1008469107. PMC 2922224. PMID 20643950.
- ^ Alliegro MC (2011). "The Nucleolinus: A disappearing, forgotten and (maybe) misnamed organelle". Communicative & Integrative Biology. 4 (2): 147–9. doi:10.4161/cib.4.2.14545. PMC 3104566. PMID 21655427.
- ^ Alliegro MC, Alliegro MA (2013). "Localization of rRNA transcribed spacer domains in the nucleolinus and maternal procentrosomes of surf clam (Spisula) oocytes". RNA Biology. 10 (3): 391–6. doi:10.4161/rna.23548. PMC 3672282. PMID 23324608.
External links edit
- Cooper, Geoffrey M. (2000). "The Nucleolus". The Cell: A Molecular Approach (2nd ed.). Sunderland MA: Sinauer Associates. ISBN 0-87893-106-6.
{{cite book}}: External link in|chapterurl=|chapterurl=ignored (|chapter-url=suggested) (help) - Nucleolus under electron microscope II at uni-mainz.de
- Nuclear Protein Database – search under compartment
- Cell+Nucleolus at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
- Histology image: 20104loa – Histology Learning System at Boston University